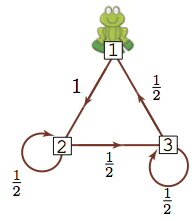
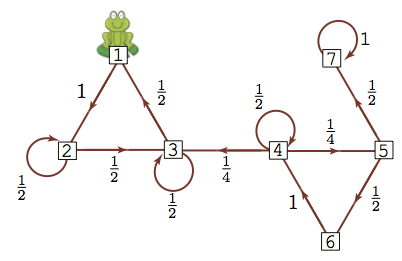
Let's do the same thing: if we start the frog at different locations, compute the probability of being at any of the lily pads after 64 steps.
## 2^6-step probabilities, starting at 1
lambda = c(1, rep(0, 6))
lambda %*% pow_P(P_big, 6)
## [,1] [,2] [,3] [,4] [,5] [,6] [,7]
## [1,] 0.2 0.4 0.4 0 0 0 0
## 2^6-step probabilities, starting at 7
lambda = c(rep(0, 6), 1)
lambda %*% pow_P(P_big, 6)
## [,1] [,2] [,3] [,4] [,5] [,6] [,7]
## [1,] 0 0 0 0 0 0 1
## 2^6-step probabilities, starting at 6
lambda = c(rep(0, 5), 1, 0)
lambda %*% pow_P(P_big, 6)
## [,1] [,2] [,3] [,4] [,5] [,6]
## [1,] 0.1333333 0.2666667 0.2666667 1.905188e-09 6.499812e-10 4.435003e-10
## [,7]
## [1,] 0.3333333
round(pow_P(P_big, 6), digits = 2)
## [,1] [,2] [,3] [,4] [,5] [,6] [,7]
## [1,] 0.20 0.40 0.40 0 0 0 0.00
## [2,] 0.20 0.40 0.40 0 0 0 0.00
## [3,] 0.20 0.40 0.40 0 0 0 0.00
## [4,] 0.13 0.27 0.27 0 0 0 0.33
## [5,] 0.07 0.13 0.13 0 0 0 0.67
## [6,] 0.13 0.27 0.27 0 0 0 0.33
## [7,] 0.00 0.00 0.00 0 0 0 1.00